


Go to chapter: 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 | 28 | 29 | 30 | 31 | 32 | 33 | 34 | 35 | 36 | 37 | 38 | 39
Chapter 3 (page 12)

Go to chapter: 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 | 28 | 29 | 30 | 31 | 32 | 33 | 34 | 35 | 36 | 37 | 38 | 39
Chapter 3 (page 12)
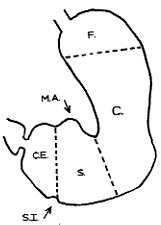 | Fig. 3.3. Muscular regions of stomach according to Forssell, F, fornix; C, corpus; S, sinus; C.E., canalis egestorius; M.A., membrana angularis; S.I., sulcus intermedius |
On the duodenal side, however, a fibrous septum separated the circular muscle of the
pylorus from the corresponding coat of the duodenum. Some of the pyloric longitudinal
fibres were continuous with those of the duodenum. The longitudinal fibres on the lesser
curvature, a short distance orally to the pyloric aperture, were weak or scanty, leading to
an intermittent outpouching of the lumen called the membrana angularis (Fig 3.3).
Forssell (l913) looked upon the canalis egestorius as an anatomically preformed structure.
On the greater curvature it extended from the sulcus intermedius to the pyloric aperture
(Fig. 3.3). The sulcus intermedius, present in anatomical specimens, was due to tonic
contraction at the entrance to the canalis, and not to an independent muscular structure.
In anatomical specimens the distance from the sulcus intermedius to the pylorus was
found to be 3.5 to 5.0 cm in non-contracted stomachs, and 3.0 to 4.5 cm in contracted
ones. On the lesser curvature Forssell's canalis included part of the membrana angularis;
he called the area between the latter and the pyloric aperture the "end piece".
The pyloric ring at the aboral end of the canalis was not a separate anatomical structure
but formed part of the musculature of the canalis. However, Forssell stated that "this
does not diminish, even in a small way, its anatomic or physiologic character as a
sphincter" and that, on account of its anatomic structure, the pyloric ring could be looked
upon as a proper sphincter, with both constrictor and dilator mechanisms.
During life the enire canalis contracted concentrically, an event which Forssell called the
"annular wave". The contraction commenced more or less in the region of the sulcus
intermedius, at a distance of 2.5 to 4.0 cm from the pylorus.
Horton (l928) studied the distribution and arrangement of the circular and longitudinal
musculature in 90 normal, fresh post-mortem stomachs; this included the study of 5171
microscopic sections. An attempt was made to determine the percentage of circular and
longitudinal fibres in the pyloric region which were continuous with the corresponding
fibres of the duodenum. The circular musculature of the pyloric canal was found to be 4
to 5 times as thick as the longitudinal; it was broken up into incomplete bundles by
connective tissue septa which entered the muscle from the submucosa and ran at right
angles to the long axis of the lumen. The septa usually extended through the circular as
far as the longitudinal layer; circular bundles formed in this way were not separate rings,
but anastomosed freely with one another.
Of 84 specimens examined, 81 showed a complete break between the circular muscle of
the pyloric region and that of the duodenum; this was confirmed in 97 percent of 1210
microscopic sections. The break between the two circular coats, caused by a septum of
connective tissue and blood vessels, was up to 1.5mm wide, but as a rule somewhat
narrower than this. A few minor variations were encountered; in 3 cases, for instance, a
small quantity (less than 2 percent) of pyloric circular fibres were carried over into the
duodenum for a short distance. The circular musculature of the duodenum, which was
much thinner than that of the pylorus, began on the distal aspect of the connective tissue
septum.
Previous Page | Table of Contents | Next Page
© Copyright PLiG 1998